
| 巹偺廋巑壽掱偵偍偗傞尋媶栚揑偼丄傛傝惛枾側怱幒嬝嵶朎丒揹婥惗棟妛儌僨儖偺峔抸丄媦傃丄怱幒嬝嫊寣忬懺偵偍偗傞僔儈儏儗乕僔儑儞夝愅偡側傢偪in silico幚尡乮僐儞僺儏乕僞忋偵峔抸偟偨儌僨儖傪梡偄偰栻暔搳梌側偳偺條乆側忦審壓偱偺怳傞晳偄傪梊應偟丄嵶朎偺婡擻傗栶妱傪暘愅偡傞幚尡乯偱偁傞丅 |
|
偼偠傔偵 怱嬝嵶朎偵偼丄枌揹埵傗嬝廂弅偲偄偭偨丄懡偔偺帠徾傪摑崌揑偵挱傔傜傟傞僔儞僾儖側弌椡偑偁傝丄傑偨僷僢僠僋儔儞僾朄偵傛傞扨暘巕偺朿戝側僨乕僞偑偁傞丅偟偐偟丄怱嬝嵶朎撪偺婡擻偲梫慺娫偺憡屳嶌梡丄怱嬝戙幱丄嫽暠楢娭偵偍偄偰丄枹偩夝柧偝傟偰偄側偄偙偲偼懡偔丄偦偺婡擻夝柧偑媮傔傜傟傞丅 嫊寣偲偼丄寣棳偺尭彮偵傛傞巁慺傗摐暘偺寚朢傪擖椡巋寖偲偟丄攺摦傗廂弅椡偺堎忢傪弌椡偲偡傞尰徾偲懆偊傜傟傞丅ATP偺嶻惗丄徚旓偺椉懁柺傪娷傓儌僨儖傪梡偄傞偙偲偱丄嵶朎儗儀儖偱偺嫊寣傪橂嵴揑偵夝愅偱偒傞偙偲偑婜懸偝傟丄嫊寣偑傕偨傜偡嵶朎撪偺婡擻傗彅梫慺偺憡娭娭學偺曄壔偵偮偄偰丄擖椡丒弌椡傪娷傔偨斖埻偱偺夝愅傪帋傒偨偄丅 怱幒嬝嵶朎偺嫊寣忬懺帪偵偍偗傞昦懺夝愅偵岦偗丄弔妛婜偼KATP偵傛傞怱嬝曐岇嶌梡偺娭傢傝偵偮偄偰僀僆儞僠儍僱儖傪拞怱偵専徹傪峴偭偨丅偦偺夁掱丄媦傃丄寢壥傛傝摼傜傟偨峫嶡偲栤戣揰偵偮偄偰偼丄屻弎偡傞丅傑偨怱幒嬝嵶朎偺嫊寣忬懺偺僔儈儏儗乕僔儑儞傪幚尰偡傞忋偱丄ATP偺摦懺傪摦揑偵儌僨儖壔偡傞偙偲偼寚偐偣側偄丅偦偙偱廐妛婜偵偼丄怱嬝嫽暠亅廂弅儌僨儖(埲壓丄 Kyoto model)[1][2]傪梡偄偰丄怱幒嬝嵶朎偺嫊寣忬懺偺僔儈儏儗乕僔儑儞傪幚尰偡傞偨傔偵丄僄僱儖僊乕戙幱宱楬偺幚憰偵庢傝慻傫偩丅Kyoto model偼丄嬝廂弅摍偺ATP徚旓偵偮偄偰惛枾側儌僨儖壔偑側偝傟偰偄傞偺偱丄徚旓傪憡曗偡傞ATP嶻惗儌僨儖偲偟偰丄巁慺傗摐暘傪庴偗庢偭偰ATP傪嶻惗偡傞丄夝摐宯丄TCA夞楬丄巁壔揑儕儞巁壔宱楬傪拞怱偲偟偨僄僱儖僊乕戙幱宱楬偺幚憰傪峴偭偨丅嵶朎枌偵傛傞婎幙偺桝憲挷愡傗丄儈僩僐儞僪儕傾枌偵傛傞娨尦摉検偺桝憲挷愡婡峔偑摑崌偝傟偰丄怱憻偼偄偮偄偐側傞忬懺偵偍偄偰傕婎幙傪帄揔偵棙梡偱偒傞傛偆偵僄僱儖僊乕嶻惗懍搙傪挷愡偡傞丅ATP嶻惗偐傜徚旓偝傟傞傑偱偺堦楢偺嶌梡偲僶儔儞僗傪E-CELL System忋偵峔抸偟丄妶摦揹埵宍惉偲偺憡娭傪専徹偟偨丅
乣KTP僠儍僱儖偺妶惈壔偲嵶朎撪梫慺偺憡娭偵娭偡傞専徹乣 怱嬝嫊寣偲偦偺敪惗婡彉 怱嬝嫊寣偲偼怱憻傪梴偆寣娗偺摦柆峝壔摍偵傛傝丄怱憻偵廫暘巁慺偑嫙媼偝傟側偄忬懺傪巜偡丅乮傛偔怱憻敪嶌偲尵偆偑丄姰慡偵摦柆偑暵嵡偟偰怱嬝偺堦晹偑夡巰偟偰偟傑偭偨応崌傪怱嬝峓嵡偲尵偄丄嫹怱徢偼偦偺慜抜奒偱偁傝丄曻偭偰偍偔偲怱嬝峓嵡偵側傞壜擻惈偑崅偄昦懺偱偁傞丅 嵶朎偺巁慺徚旓検偑憹壛偡傞偵偮傟偰丄侾屄偺怱嬝嵶朎偺拞怱晹偵巁慺晄懌椞堟偑偱偒丄偦傟偑儈僩僐儞僪儕傾偺戙幱傪梷惂偡傞丅怱嬝偱偼丄巁慺暘巕偼嵶朎昞柺偐傜嵶朎拞怱傑偱栺10儈僋儘儞偺嫍棧傪奼嶶偡傟偽傛偄偺偩偑丄偙偺嫍棧偼丄巁慺暘巕偵偲偭偰挿偄摴偺傝偱偁傝丄廬偭偰丄摿偵偙偺摴偺傝偺廔拝揰偱偁傞嵶朎拞怱晹偵埵抲偡傞儈僩僐儞僪儕傾偱偼丄巁慺廀梫偑崅傑傞偲丄奼嶶偵傛傞巁慺嫙媼偑捛偄偮偐側偔側偭偰偟傑偆偺偱偁傞丅
峫嶡 (1) 嘆 ATP擹搙尭彮偵敽偄丄KATP僠儍僱儖偼妶惈壔偟丄妶摦揹埵偑抁弅偡傞偙偲偑妋擣偝傟偨丅偙傟偼怱嬝嵶朎撪偺俙俿俹晄懌偵傛傝丄KATP僠儍僱儖偑妶惈壔偝傟丄怱嬝偺妶摦揹埵偺帩懕帪娫傪抁弅偟偨偲峫偊傞丅 嘇 揹埵埶懚惈僠儍僱儖偱偁傞丄ICaL丄INa丄IK倰丄IK倱丄IRK1丄ICaT丄Na+/K+ pump丄Na+/Ca2+Exchanger偼偡傋偰丄戞俀乣俁憡晅嬤偱偺揹棳敪惗帪娫暆偺抁弅偑尒傜傟偨丅偙傟偼枌揹埵偵傛偭偰僼傿乕僪僶僢僋偑偐偐偭偨偲峫偊傞丅 嘊 Na+/K+ pump傕Na+/Ca2+ Exchanger傕ATP偵埶懚偟偨挷愡婡擻傪傕偮丅崱夞丄ATP擹搙偺尭彮偵敽偆揹棳検偺憹壛傪妋擣偡傞偙偲偑偱偒偨丅 嘋 堦曽丄SR Ca2+Uptake偼壖愢偵斀偟丄ATP擹搙偺尭彮偵敽偄丄揹棳検偑憹壛偡傞偙偲偼側偔丄傓偟傠偽傜偮偒偑尒偊偨丅妶摦揹埵傪抁偔偡傞偙偲偱Ca2+偺棳擖傪梷偊丄俙俿俹偺徚旓傪嬌椡梷偊傛偆偲偡傞怱嬝曐岇嶌梡偑摥偄偰偄傞偲峫偊傜傟傞丅偟偐偟丄嵶朎撪偺Ca2+擹搙偺梷惂傗APD傗KATP僠儍僱儖偺妶惈壔偲偺憡娭偑尒傜傟側偐偭偨丅 乣儈僩僐儞僪儕傾丒僄僱儖僊乕戙幱宱楬儌僨儖偺幚憰乣 怱嬝嵶朎偺儈僩僐儞僪儕傾偵偮偄偰 儈僩僐儞僪儕傾偺嵟戝偺婡擻偼丄揹巕揱払宯偵傛傝儈僩僐儞僪儕傾枌娫峯乮撪枌丒奜枌娫乯偺僾儘僩儞擹搙傪崅傔丄擹搙岡攝偵傛傞儈僩僐儞僪儕傾撪傊偺僾儘僩儞僀僆儞偺攔弌傪嬱摦椡偲偟偰丄ATP崌惉峺慺偵傛傝ATP傪嶻惗偡傞偙偲偱偁傞丅 儈僩僐儞僪儕傾偵傛傞Ca2+庢傝崬傒偼丄撪晹偑晧偵暘嬌偟偰偄傞儈僩僐儞僪儕傾枌揹埵偵埶懚偟偰偍傝丄偙傟傪峴偆暘巕偼mitochondrial Ca2+ uniporter (mCU)偲屇偽傟偰偄傞丅堦曽丄Na+埶懚惈偵儈僩僐儞僪儕傾偐傜Ca2+傪攔弌偡傞抈敀傕偁傝丄偙傟偼mitochondrial Na+/Ca2+exchanger (mNCE)偲偄偆丅傕偆堦偮丄mitochondrial permeability transition pore (mPTP)偲屇偽傟傞摟夁惈偺崅偄僠儍僱儖偑偁傞丅mPTP偼惗棟揑忦審壓偱偼奐岥偟側偄偙偲偼傢偐偭偰偄傞偑丄枹偩夝柧偝傟偰偄側偄揰傕懡偄丅偦偺懠偵丄Rapid uptake mode偲屇偽傟傞丄嵶朎幙偺Ca2+擹搙偺怳摦偵墳偠偰摥偔僩儔儞僗億乕僞乕偑偁傞丅傑偨丄Na+旕埶懚惈偺Ca2+攔弌婡峔偑偁傞偲偺愢傕偁傞丅
 Table1: Steady State Parameters
2003擭搙妛夛敪昞 [1]Miyamoto, S., Naito, Y., Yoneda, M., Ishinabe, S., Sarai, N., Matsuoka, S., Noma, A., Tomita, M., Electrophysiological Simulation of Ventricular Cell Using E-CELL System and in preparation for construction ischemic state model., First IECA Conference on Systems Biology of E.coli(2003 Tsuruoka) [2]Motohiro Yoneda, Shoko Miyamoto, Yasuhiro Naito, Sayaka Ishinabe, Nobuaki Sarai, Satoshi Matsuoka, Akinori Noma, Masaru TomitaModel Construction of the Energy Metabolism Pathway in Myocardial Cell, International Conference On Genome Informatics.(2003 Yokohama) [3]Miyamoto, S., Naito, Y., Yoneda, M., Ishinabe, S., Sarai, N., Matsuoka, S., Noma, A., Tomita, M., 嫊寣忬懺偺僔儈儏儗乕僔儑儞偵岦偗偨怱幒嬝嵶朎儌僨儖偺奼挘., 戞26夞擔杮暘巕惗暔妛夛擭夛.(2003) |
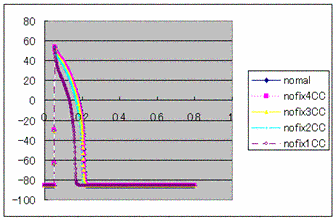 Figure1:
Membrane potential by CC method
Figure1:
Membrane potential by CC method 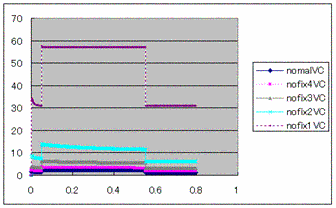 Figure2: The
current of KATP channel by VC method
Figure2: The
current of KATP channel by VC method  Figure3: The
current of Na+/K+ pump by VC
method
Figure3: The
current of Na+/K+ pump by VC
method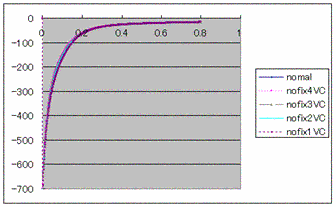 Figure4: The
current of SR Ca2+Uptake by VC method
Figure4: The
current of SR Ca2+Uptake by VC method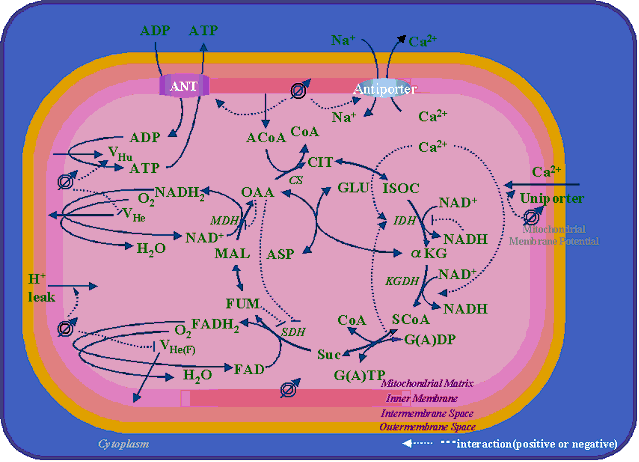 Figure5:
儌僨儖偵慻傒崬傑傟偰偄傞揹婥惗棟妛揑側儈僩僐儞僪儕傾偺戙幱嶌梡偺夁掱丄媦傃偦傟傜偺憡屳嶌梡偺峔憿恾
Figure5:
儌僨儖偵慻傒崬傑傟偰偄傞揹婥惗棟妛揑側儈僩僐儞僪儕傾偺戙幱嶌梡偺夁掱丄媦傃偦傟傜偺憡屳嶌梡偺峔憿恾